


We aimed to investigate the effects of AR-A014418, a strong inhibitor specific to GSK-3beta, on neuronal apoptosis and neuroprotection in the traumatic SCI model.
Materials and methodsIn this study, three groups were generated from 36 Wistar rats; (1) control, (2) spinal cord trauma group created by clip compression technique after laminectomy, and (3) AR-A014418 (4mg/kg, i.p., DMSO) treatment group after laminectomy and spinal cord trauma. The TUNEL assay for apoptosis detection, immunohistochemical staining for bax and TGF-beta were applied in spinal cord tissues. For light microscopic examination, necrotic, and apoptotic cells were counted, and PMNL counting was applied to detect inflammation. Functional recovery was tested by field locomotor test in the 3rd and 7th days following surgery.
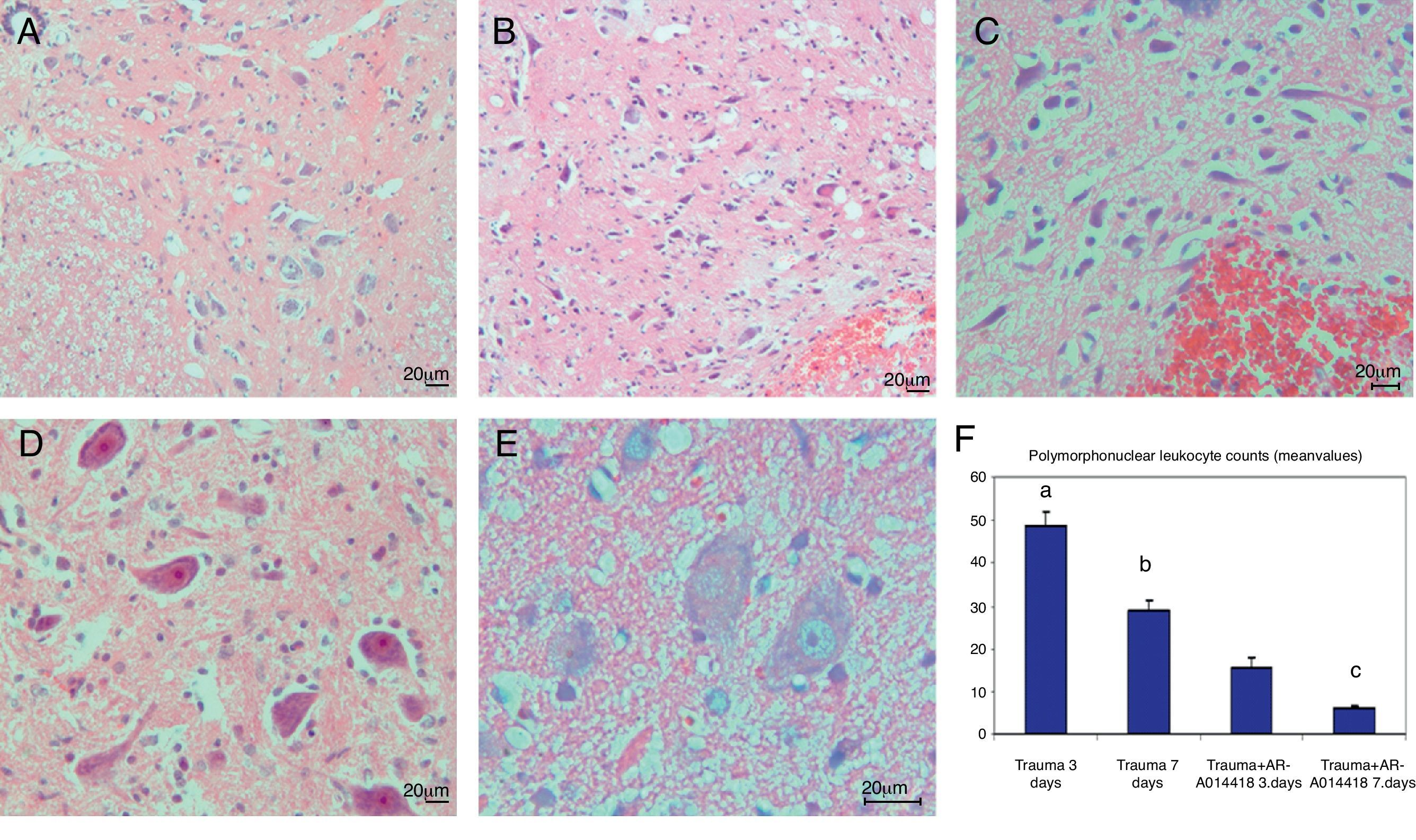
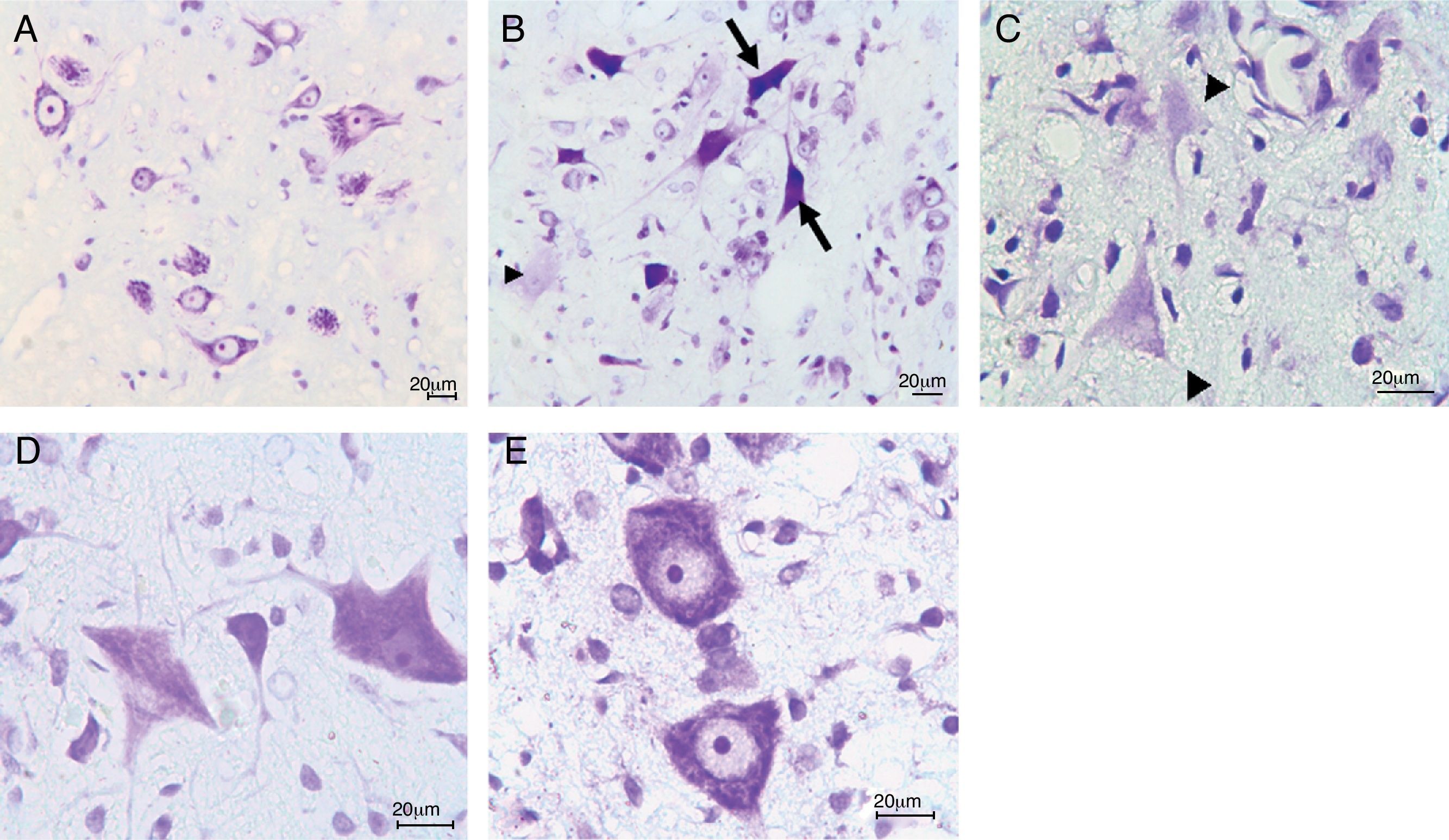
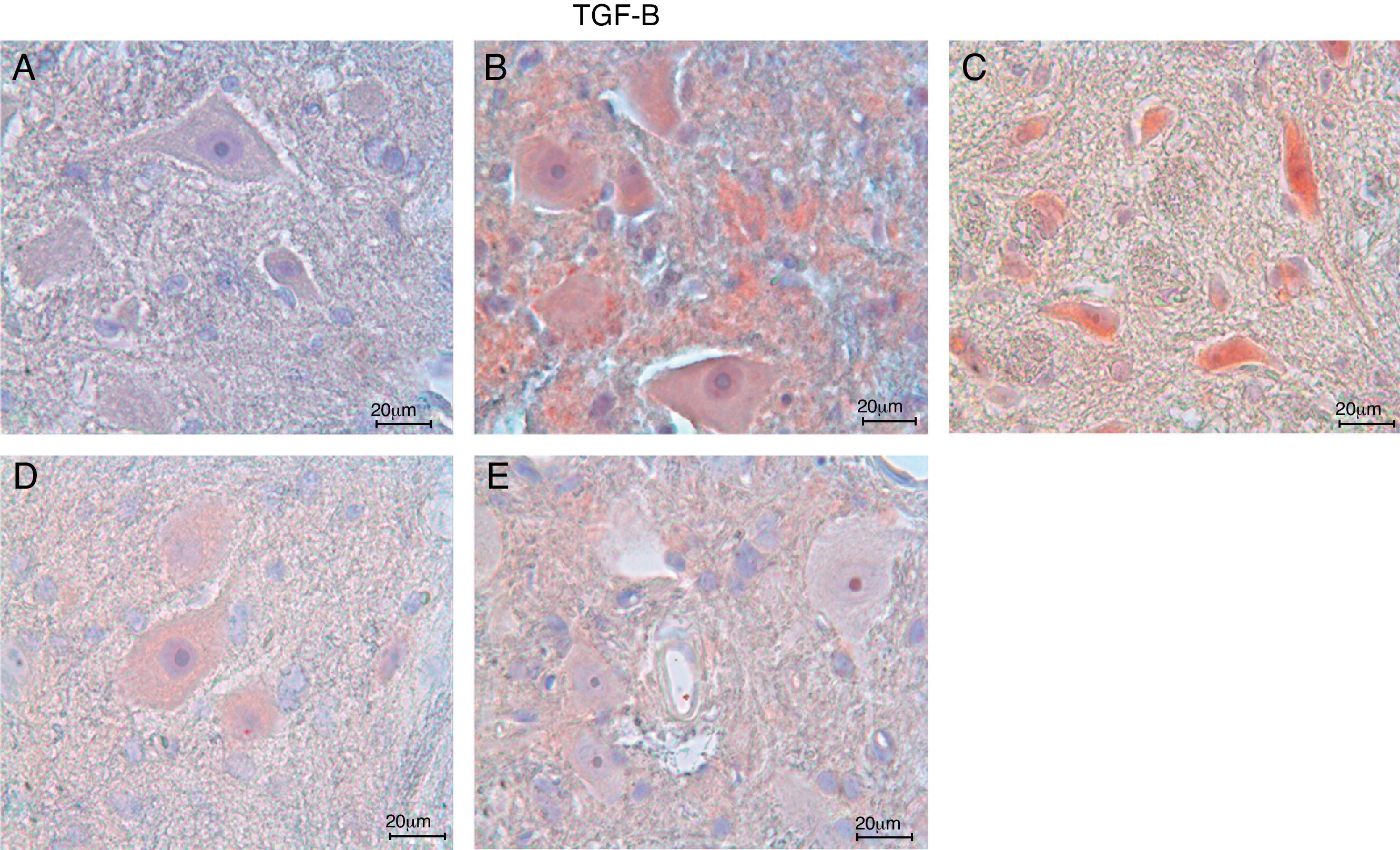
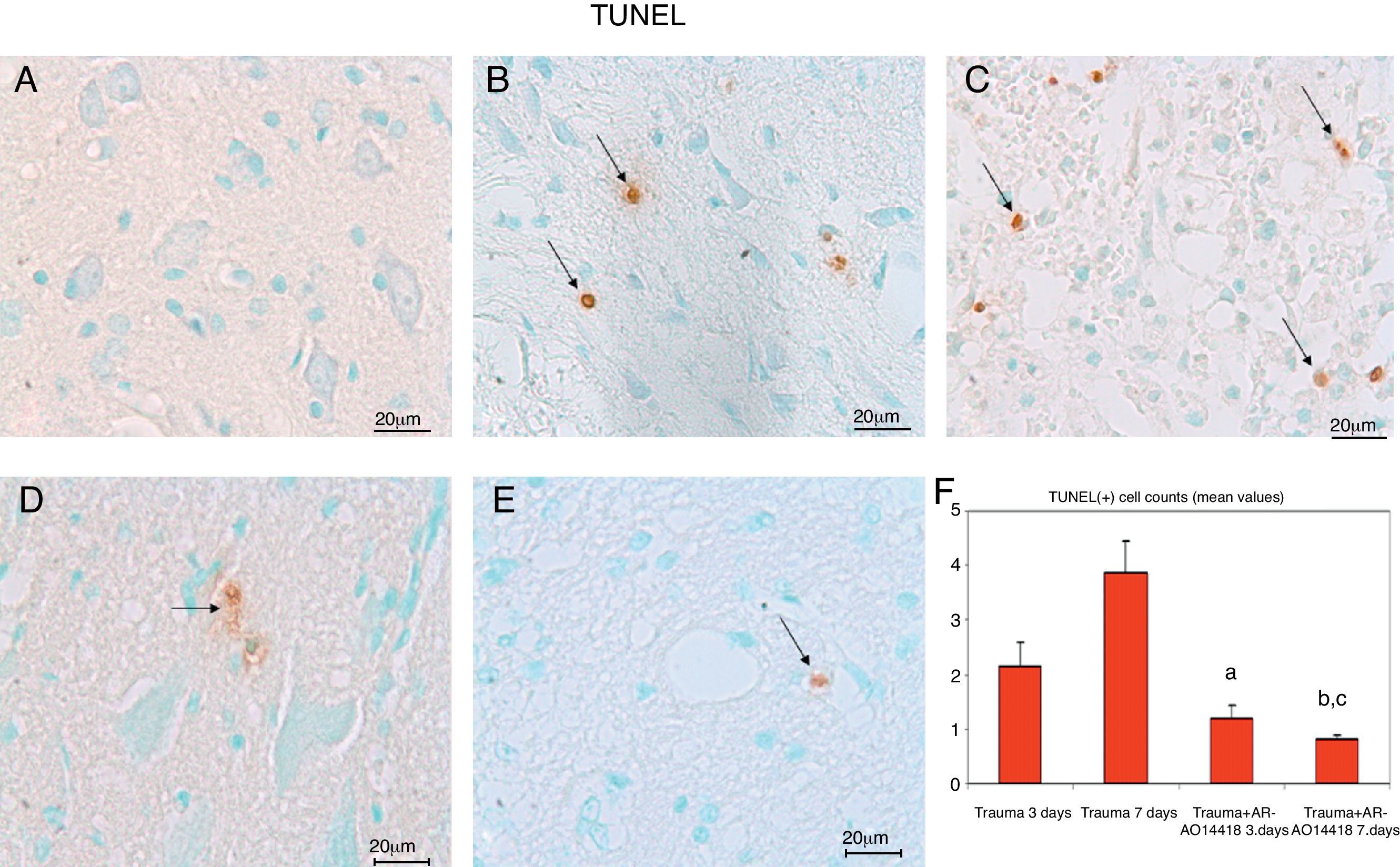
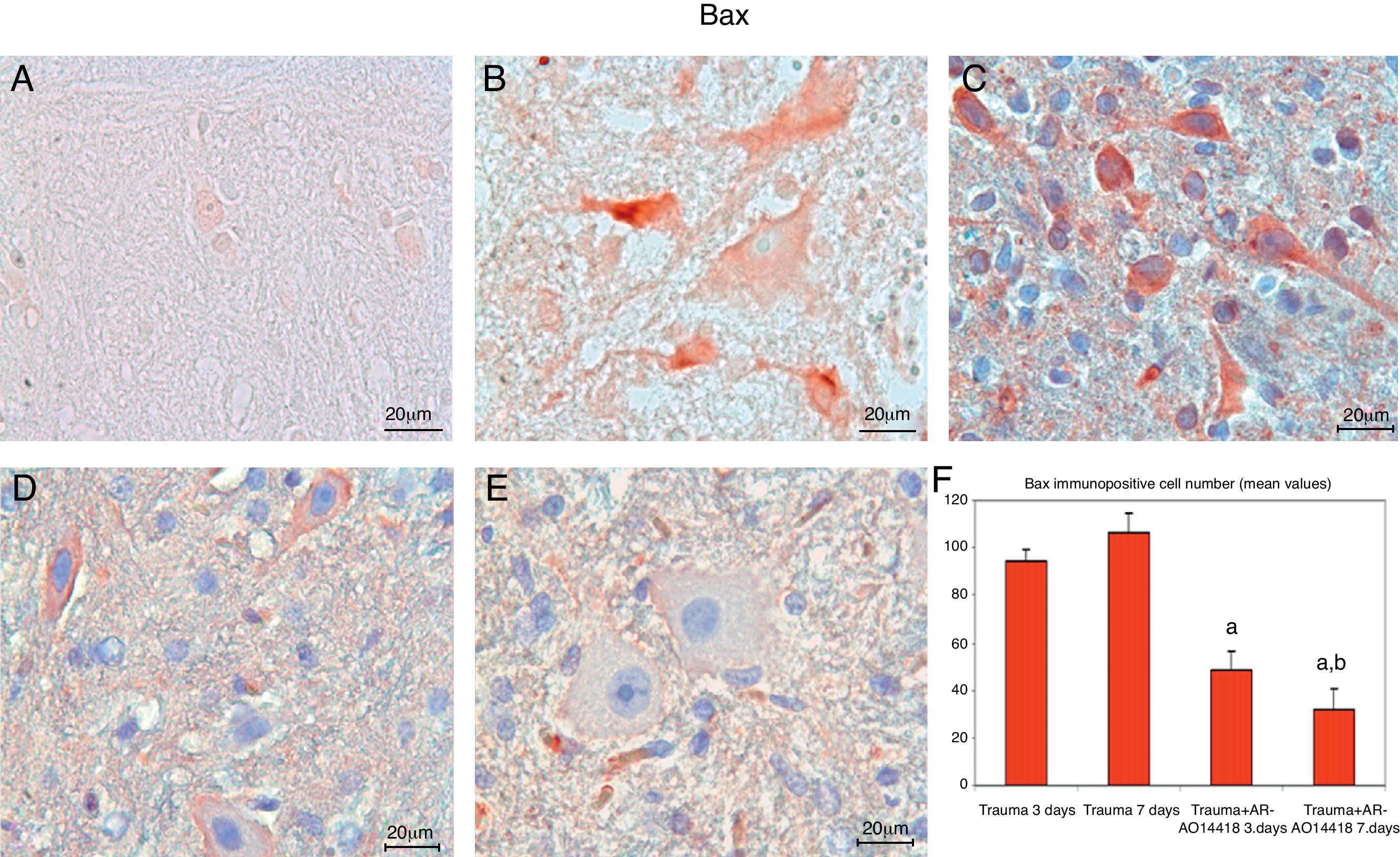
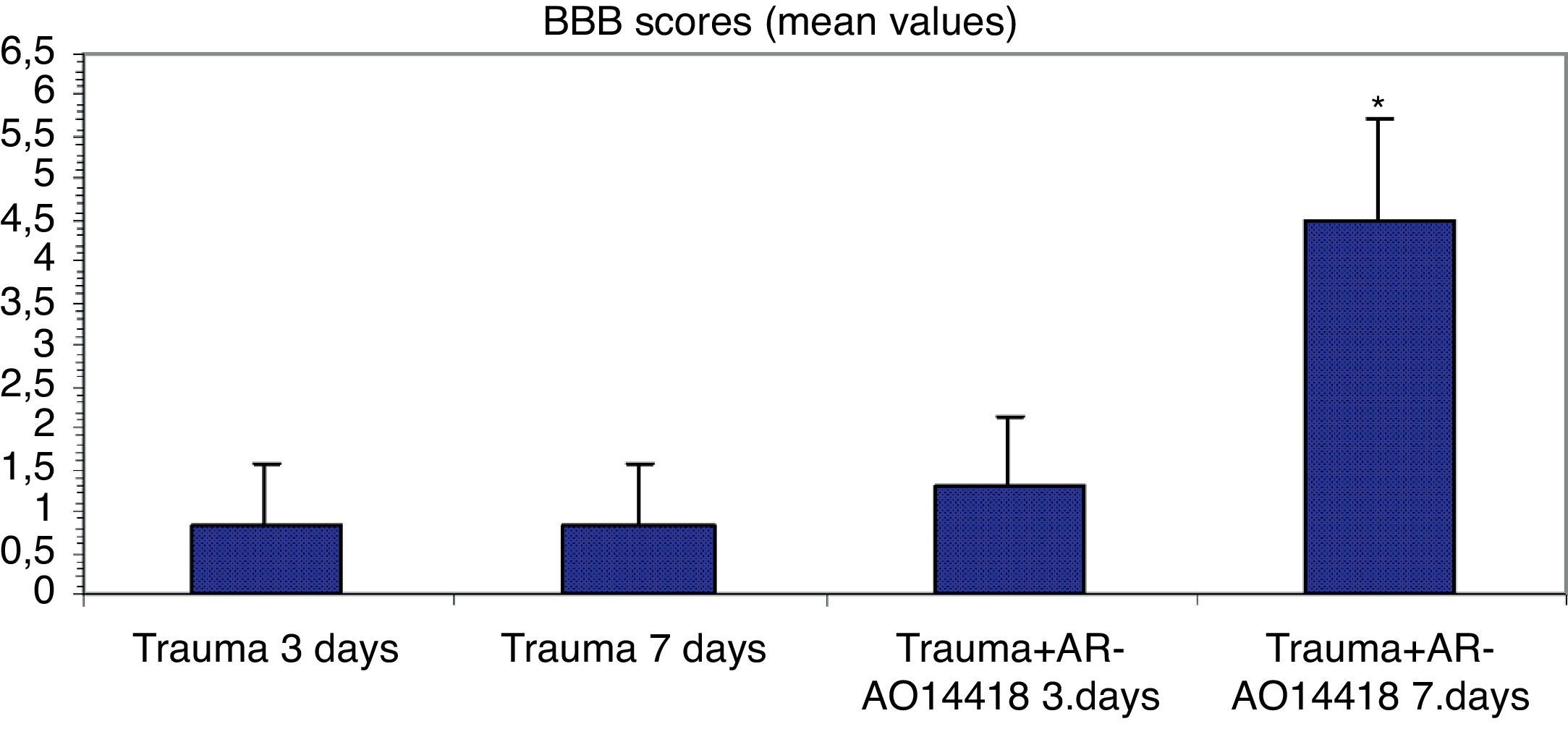
ResultsIn the trauma group, diffuse hemorrhage, cavitation, necrosis and edematous regions, degeneration in motor neurons and leukocyte infiltration were observed in gray matter. In the AR-A014418-treated groups, healthy cells were observed in more places compared to the trauma groups, however, cavitation, hemorrhagic, and edematous areas were seen in gray matter. In the AR-A014418-treatment groups, the number of apoptotic cells in the 3rd and 7th days (respectively; p<0.05, p<0.01), were significantly decreased compared to the trauma groups, as were the levels of bax (p<0.01) and TGF-beta 1 immunoreactivity. Results of the locomotor test were significantly increased in the treatment group (p<0.001) as compared to the trauma group.
ConclusionsIn this experimental spinal cord trauma model study neural apoptosis was significantly triggered in secondary damage developed after trauma, however, neurological healing was expedited by preventing mitochondrial apoptosis and reducing the inflammation by the potent inhibitor AR-A014418, which is GSK-3beta selective.
Este trabajo tuvo como objetivo investigar los efectos del AR-A014418, un potente inhibidor específico de la GSQ-3beta, en la apoptosis y neuroprotección neuronales en el modelo de lesión medular traumática.
Materiales y métodosSe generaron tres grupos a partir de 36 ratas Wistar: (1) control, (2) grupo de traumatismo medular obtenido mediante técnica de pinzamiento post-laminectomía, y (3) grupo de tratamiento mediante AR-A014418 (4mg/kg, i.v., DMSO) post-laminectomía y traumatismo medular. Se aplicaron el test TUNEL para la detección de la apoptosis, tinción inmunohistoquímica para bax y TGF-beta en los tejidos medulares. Se llevo a cabo un examen microscópico y recuento de células necróticas y apoptóticas, así como recuento de LPMN para detectar inflamación. La recuperación funcional fue verificada mediante la prueba de campo del aparato locomotor en los días 3 y 7 después de la cirugía.
ResultadosSe observo hemorragia difusa, cavitación, necrosis y regiones edematosas, degeneración de las neuronas motoras e infiltración leucocítica en la materia gris en los grupos traumáticos. En los grupos con tratamiento AR-A014418 se observaron células sanas con mayor abundancia en comparación con los grupos traumáticos. Sin embargo, se observaron áreas de cavitación, hemorragia y edema en la materia gris. En los grupos con tratamiento AR-A014418, el número de células apoptóticas en los días 3 y 7 (p<0,05 y p<0,01, respectivamente), se redujo significativamente en comparación con los grupos traumáticos, así como los niveles de bax (p<0,01) y las células inmunopositivas para TGF-beta 1. Los resultados de la prueba del aparato locomotor se incrementaron significativamente en el grupo de tratamiento (p<0,001) en comparación con el grupo traumático.
ConclusionesMediante un estudio experimental de un modelo de traumatismo medular se observó como la apoptosis neuronal era desencadenada de manera significativa por daños secundarios postraumáticos. Sin embargo, la mejora neurológica se vio acelerada por la prevención de la apoptosis mitocondrial y reducción de la inflamación causadas por el inhibidor de GSQ-3beta, AR-A014418.
Article
![]()
If it is the first time you have accessed you can obtain your credentials by contacting Elsevier Spain in suscripciones@elsevier.com or by calling our Customer Service at902 88 87 40 if you are calling from Spain or at +34 932 418 800 (from 9 to 18h., GMT + 1) if you are calling outside of Spain.
If you already have your login data, please click here .
If you have forgotten your password you can you can recover it by clicking here and selecting the option ¿I have forgotten my password¿.



